抑制性神経
関連概念 : 生物学
[inhibitory neurons]
抑制性神経細胞。抑制性ニューロン。
神経細胞の一で、抑制性シナプスを形成する(A. Interneurons unbound; Klausberger&Somogyi,2008)。
介在神経の大部分を占める。
【新皮質抑制性細胞】
◆神経伝達物質
・GABA作動性神経
・グリシン作動性神経
・GABA・グリシン作動性神経(Ornung et al.,1996)
◆形態
Patilla terminology(Petilla Interneuron Nomenclature Group,2008)
(Somogyi,Salient,1998; DeFelipe,1997; Markram,Wu,2004)
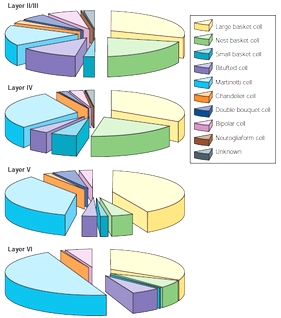
○籠状細胞(抑制性神経の50%)soma-targeting
錐体細胞や他の介在神経のsomaや近位dendに出力
▲large basket cells (LBCs)(Kisvarday&Eysel,1992)
古典的な籠状細胞で広くaxon分岐し、近傍や離れたコラムも抑制
(層内コラム間の側抑制の主成分)
普通depressing synapses
▲small basket cells (SBCs)(Kisvarday,Somogyi,1985)
soma-targetingで局所的に密でhighly varicoseなaxon分岐
錐体細胞でのシナプス入力のうち最も頻繁
・the clutch cell(視覚野IV層のSBCの一種)
中型で軸索collateralsに大きいbulbous terminals
▲nest basket cells (NBCs)
LBCとSBCの間のようだが局所高密度軸索はSBCに類似
○シャンデリア細胞(Chandelier cells)axon-targeting
樹状突起統合後のspikeを最終的に抑制可能
(Buhl,Somogyi,1994; Miles,Hajos,1996; Zhu,Zhu,2004)
II-VI層や海馬(Ganter,Somogyi,2004)に分布
○dendritic-targeting interneurons
(proximal dendrite-targeting, dendrite-targeting
dendriteand tuft-targeting)
▲マルティノッティ細胞(Martinotti cells)
II-VI層に分布し、I層で軸索を水平にmm単位で広げる
錐体細胞tuft樹状突起を抑制(DeFelipe,2002)
・Infragranular MCsはIV層へ選択的に投射
▲皮質双極細胞(BPCs)VIP放出する興奮性細胞もある
小さい細胞でspindle or ovoid somataを持ちII-VI層に分布
樹状突起をI-V層に伸ばし狭い帯(<50um)の軸索で全層抑制
(Peters&Harriman,1988; Peters,1990)
数個の錐体細胞basal dendに出力
▲ダブルブーケ細胞(Double bouquet cells)
II-V層に分布して緊密な線維性axonal cylinder形成
(Somogyi&Cowey,1981; DeFelipe,Jones,1990)
varicoseなcollateralsを全層に伸ばしbasal dend抑制
▲皮質双房細胞(Bitufted cells)
ovoid somaで垂直両極にdend伸ばしてII-VI層に存在
軸索は垂直には短く水平に広く伸ばしコラム超えることも
▲ニューログリアフォーム細胞(Neurogliaform cells)
小型でbutton-type細胞で多くの細い放射状dendを短く伸ばす
非常に細い軸索をsomaかdend基部から出しすぐに密に分岐
数十個に分岐してfine boutons持つ
▲皮質1層介在神経(Layer I interneurons)
・Cajal Retzius cells
大きい細胞で水平突起持ちI層でのみ主に発生途中に存在
・heterogeneous group of small,multipolar interneurons
垂直に軸索伸ばす(poor and rich axonal plexus cells)
◆発火特性での分類
onsetとsteady-state firingで分類
(Connors&Gutnick,1990; Kawaguchi,1993)
(Gupta,Markram,2000)
○non-accommodating (NAC)I-VI層
発火率は電流に対して急峻に増大して順応ほとんどしない
APは短く深く速いAHP(fAHP)
juvenile somatosensory cortexでは最も多い
・bursting NAC(fast spiking(FS))onsetで>3AP

・delay NAC(FS, LS)
Late-spiking (LS) cells(Kawaguchi&Kubota,1998)
脱分極させたとき発火まで潜時(II/III層とV層)
いくつかはNGCs

・classical NAC(FS)onsetでburstingやdelayない

○accommodating (AC)II-VI層
juvenile somatosensory cortexでは最も多い
・bursting AC(BSNP: burst-spiking non-pyramidal)
low-threshold-spiking (LTS) cells
(burst-spiking non-pyramidal cells: BSNP cels)
(Kawaguchi&Kubota,1993;1997; Kawaguchi,1993; )
V層に存在し過分極後にバースト状発火し一部はMCs,DBCs
V層錐体細胞もしばしばburst

・delay AC(FS, LS)

・classical AC(RSNP)
Regular-spiking non-pyramidal (RSNP) cells
錐体細胞に発火類似(Kawaguchi&Kubota,1996; 1998)
II/III層やV層に存在。一部はMCs, DBCs, BPCs

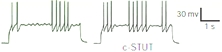
○stuttering (STUT)II-VI層
様々な電流に対し高頻度burstと不規則なsilence(Morsecode-like)
juvenile somatosensory cortexではACやNACよりは少ない
・bursting STUT(BSNP)

・delay STUT(FS, LS)

・classical STUT(FS)
○irregular spiking (IS)II-V層
1APを不規則に生じる
数は少なく形態bipolarでII/III層やV層に分布
・bursting IS(電流注入直後のみにburst)

(Porter,Audinat,1998)
・IS1 cells(burst短い)
・IS2 cells(burst長い)
・delay IS
・classical IS

○bursting (BST)II-V層(特にV層に多い)
遅い脱分極で3-5 APsのburstした後、遅いAHP(sAHP)
・repetitive BST burst発火繰り返し(chattering様)

・transient BST(BSNP)強いburstをonsetのみ

・initial BST(BSNP)初めburstするが次にゆっくりとした発火

◆遺伝子・タンパク質による分類
(Toledo-Rodriguez,Markram,2004)
(Cauli,1997; DeFelipe,1993)
○CBPによる分類
(Demeulemeester,Pochet,1989; Rogers&Resibois,1992;Cauli,2000)
概ね上手く分かれるが時に複数のCBP発現(特にCBとPV、またCBとCR)
▲parvalbumin陽性細胞(しばしばFS細胞: Chow,1999)
籠状細胞の半分くらいとシャンデリア細胞(DeFelipe,1999)
・HCN1, HCN2
・Kv3.1, Kv1.2, Kv1.6, Kv1.1, Kv3.2, Kvβ1
強力な遅延整流性電位依存性Kチャネル(Kv3.1,Kv3.2)
(Rudy&McBain,2001; Martina,Jonas,1998;
Erisir,Leonard,1999)
Kv3.1はPV陰性細胞や錐体細胞にも発現
遅延整流性Kv1.6も発現
修飾サブユニットKvβ1はHCN1,HCN2,Kv1の遅延整流性をA型に変化
・Caα1A
高閾値Caα1Gやその修飾サブユニットCaβ4も発現
・calretinin, Caα1I, Kv2.2, HCN4, SK2は発現少ない
▲calretinin陽性細胞(しばしばAC,IS: Vergara,Adelman,1998)
・SK2, Kv3.4
・Caα1B
▲calbindin陽性細胞(bursting特性: Ertel&Ertel,1997)
・HCN3, HCN4
・Kv1.4, Kv3.3
・Caβ4, Caα1G, Caβ1, Caβ3
・busting onset細胞とdelay onset細胞で発現ほぼ真逆
(Toledo-Rodriguez,Markram,2004)
同じFC細胞なので少ない転写因子でチャネルセット調節か
◆神経ペプチド
(Hendry,Emson,1984; Rogers,1992
Demeulemeester,Vanderhaeghen,1988; Morrison,Bloom,1984
Jones&Hendry,1986; Somogyi,1984; Hendry,1984)
・parvalbumin, calretinin, somatostatin発現細胞
(Kubota,Yui,1994)
・タンパク質レベルではPV, SOM, VIP発現細胞は異なる細胞
(Kawaguchi&Kubota,1997)
mRNAレベルでは分かれない
○somatostatin発現細胞
マルティノッティ細胞に発現(Wahle,1993)
BTCsにも発現(Meinecke&Peters,1986)
○VIP陽性細胞
マルティノッティ細胞は発現しない
small籠状細胞に発現(Valverde,1986)
DBCに発現(Kawaguchi&Kubota,1997)
BPCに発現(Demeulemeester,Vanderhaeghen,1988)
○cholecystokinin陽性細胞
【特徴】
◆分布
・新皮質の約20-30%のほとんど
◆多様
(Cauli,1997;2002; DeFelipe,1993; Kawaguchi&Kubota,1997)
(Somogyi,Buhl,1998; Gupta,Markram,2000; Thomson&Deuchars,1994)
◆形態
・小さい
・aspiny dendrites
・somaに興奮・抑制シナプス入りうる
・軸索は通常コラム内
コラムを超えるときは側方向が多い
・樹状突起の形態は多様だが、これでは分類難しい
(multipolar, bipolar, bitufted)
◆可能な神経回路
・intralaminar-interlaminar(same layer-cross layer)
・intracolumnar-intercolumnar(same column-cross column)
・neocortical/subcortical/opposite hemisphere regions
・電気シナプス
◆interneuronsの方が発火率2-3倍速い
(McCormick,Prince,1985)
◆神経伝達物質
GABA (GADで生成)
細胞質のGABA/gly濃度5-7mM程度(Apostolides&Trussell,2013)
glu濃度増大させると1min程度でGABA/gly出力増強
活動依存的に発達でグリシンへの変化
上オリーブ核(Nabekura,Ishibashi,2004; Kotak,Sanes,1998)
◆興奮性神経とのbalance
・感覚入力で興奮性neuronと同時に活動上昇
(Borg-Graham,Fregnac,1998; Monier,Fregnac,2003)
(Wehr&Zador,2003)
・時間的応答が様々なので一過性にはバランス崩れる
(Hirsch,Martinez,1998; Anderson,Ferster,2000; Tan,Schreiner,2004)
・機能
・両方向に振れられるように正規化
◆synaptic dynamics(Silberberg,Markram,2004)
◆発火特性
・形態とは必ずしも対応しない
(Gupta,Markram,2000; Wang,Markram,2002)
(Kawaguchi,1993;1995; Kawaguchi&Kubota,1997)
・in vivoとin vitroの違いも(シナプス数やneuromodulation)
・主に発現チャネルが決定(Llinas,1988; Toledo-Rodriguez,2004)
・形態による受動的特性も多少影響(Mainen,Sejnowski,1996)
◆可塑性
LTP, LTD(Nugent et al., 2007)
エンドカンナビノイド受容体(Chevaleyre et al., 2006)
preの興奮性シナプス(Kullmann and Lamsa, 2007)
◆マーカー
・mDlx(Dimidschstein,Fishell,2017) enhancer (mI56i)
distal-less homeobox 5 and 6 (Dlx5/6) が共有する400bp
Dlx5/6は胚発達時にすべての前脳GABA性神経で発現
【機能】
◆行動
・AIYの活動パターンでC. elegansに2種類の行動(Kocabas,Ramanathan,2012)
2014/04/27 masashi tanaka
