V1
関連概念 : 生物学
[V1, visual area 1; striate cortex]
17野。V1野。有線野。第一次視覚野。VI。
視覚野の一。
サルでは後頭葉に大きく露出し大脳皮質の17%を占めるが、ヒトでは主要な部分が大脳半球内側面の鳥距裂溝溝壁に位置し、大脳皮質のわずか1.5%程度である。
厚さ1.5mm程度で、顆粒細胞が密に分布する4層が発達している。
【神経連絡】
◆入力
・同側の外側膝状体背側核(視放線)
4Cに入力(ただしK層は1層や2,3層に入力)
・その他
V1神経の発火パターンを調節(McCormick,1993)
・前脳基底部からのアセチルコリン入力
マイネルト基底核など
・青斑核からのノルアドレナリン入力
・縫線核からのセロトニン入力
◆出力
・V2
・V3
・MT野
【構造】
◆超コラム(hypercolumn)(Hubel,1988)
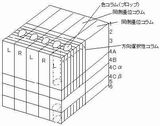
○眼優位性コラム(4層で顕著。2,3,5,6層では両眼反応もある)
幅400-500μm
ネコではサルよりも両眼応答の傾向強い
オートラジオグラフィーで発見
トレーサー(2-デオキシグルコース)を注射
活動した神経細胞は2-DGを取り込む
2-DG-6リン酸で代謝が止まる
片眼にWGA-HRP打っても見える
片眼遮蔽でコラム構造が乱れる
皮質5層の膜興奮性の変化(NatarajTurrigiano,2010)
○方位選択性コラム(2,3,4A層で顕著)(Cheng,Tanaka,2001)
水平方向:方位が10度/50μmで変化する
垂直方向:ほぼ方位が同じ
・高空間周波数コラム(特異点の周辺)
・視差選択性細胞(特異点の周辺)
光学的計測では空間分布はパッチ状(Bonhoeffer&Grinvald,1991)
水平縞のみの環境では垂直方位への応答細胞欠損(blakemore&Cooper,1970)
機序
・反回性ネットワーク(Ringach,1997; Sompolinski,1997)
刺激から40-60ms後に方位選択性最大
その後、選択的方位への応答が弱くなり、他の方位への反応が増強
・不適当方位を抑制(bicucullineで全方位に応答)
(Sillito,1975)
・抑制は最適方位以外では生じない(Ferster,1986)
・LGNからの入力が特定の傾き(Chapman,Stryker,1991)
受容野の縦横比1.7程度なので無理(Pei,1994)
○方向選択性コラム(Tootell,1981; 1983)
ブロッブ領域とその直下は、低周波数(1-1.5c/deg)に応答
ブロッブ領域は色
ブロッブ間領域とその下は、高周波数(5-7c/deg)に応答
ブロッブ間領域は形・奥行き・色
4Cβ層は4Cα層よりも高周波数に応答
ネコは霊長類に比べて低周波数(識別能力低い)
◆機能コラム
(Blasdel,1992; Bardfeld&Grinvald,1992; Shmmuel&Grinvald,1996)
○線形表現領域(方位コラム)
○ブロッブ領域
○ピンホイール(ブロッブ間領域)
特異点を中心に方位選択性細胞が放射状に配置
・低空間周波数コラム(特異点の近傍)
◆6層構造
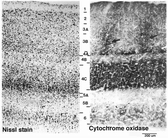
○1層:外側膝状体背側核のK層から直接投射(Lachica,1992)
○2層,3層
・ブロッブ領域
BD系(blob-dominated stream)の始点
外側膝状体背側核K層から直接入力しV2狭線条部へ
Caイメージングではblobごとに色選択性異なる(Ohki, 宮川研)
・ブロッブ間領域
ID系(interblob-dominated stream)の始点
外側膝状体背側核P層から間接的に入力しV2淡線条へ
・L2/3錐体細胞
同じ方位/方向選択性を受ける細胞から入力強い(Cossell,Mrsic-Flogel,2015)
同じ応答特性と異なる応答特性のL4,L5細胞から入力受ける細胞存在
(Wertz,Roska,2015)
一樹状突起に異なる方位/方向選択性入力(mice: Jia,Konnerth,2010)
安定spinesの方位選択性は数週間安定(Chen,Kim,2013)
spines応答の線形加算でsoma応答説明可能
PV陽性細胞でもspines方位選択性あり
樹状突起スパイクが方位選択性増強(Smith,Hausser,2013)
○4層:眼優位性カラムが顕著(ネコでは上下2亜層)
・4A 層:4Cβ層から入力?
・4B 層:運動。V3、V2広線条部へ
ベイラージャーの外線条(ジェンナリの線条)が見える
4Cαから入力
MD系(magono-dominated stream)の始点
V2広線条、V3、MT野へ出力
・4Cα層:外側膝状体背側核のM層から
4B層に出力
・4Cβ層:外側膝状体背側核のP層から
4Cは横の連絡線維が非常に発達しているため線状に見える
○5層
眼球運動の制御に関係
2,3層から入力し、上丘・視床枕などへ出力
○6層
浅層や5層から入力を受け、外側膝状体へ出力
外側膝状体の同期発火に関与(Sillito,1994)
受容野は小さく、フィードバック先の受容野と対応(注意と関係?)
ゲインコントロール(Olsen,Scanziani,2012)
end inhibition(Bolz&Gilbert,1986)
ある大きさ以上のバーに応答しない
4A,4B層あたりには出力細胞である錐体細胞があり、4C層あたりにはインターニューロンである星状細胞Stellate cellがある。
【細胞】
◆形態による分類
○有棘細胞 spiny(興奮性が多い)
・顆粒細胞(granule 星状細胞 stellate)約10%
外側膝状体から興奮性入力を受けて錐体細胞に出力
単純型細胞が多い
・錐体細胞(pyramidal)約70%
4C層以外に分布して他の大脳皮質や外側膝状体や上丘に出力
複雑型細胞や超複雑型細胞が含まれる
○無棘細胞 smooth(抑制性が多い)約20%(Szentagothai,1978)
閾値以上の脱分極に対して持続的に発火(McCormick,1985)
・大バスケット細胞
1mm以上も軸索を広げて広範囲に抑制
・バスケット細胞
単純型や複雑型細胞
・シャンデリア細胞
・クラッチ細胞
◆機能による分類
・方位選択性細胞(Nicholls et al., 2001)
同方位の線分に選択性を示す細胞同士に相関(Kapadia,1995)
図と地に属する場合で応答が異なる(Lamme&Spekreijse,2000)
図だと持続的応答
地だと一過性応答
麻酔ではこの効果がないため中枢からのfeedbackか
・ブロッブ細胞(Livingstone&Hubel)
・色選択性細胞(COブロッブ領域)
方位選択性なし
70%は波長選択性あり、30%は選択性なし
低空間周波数選択性
チトクロームオキシダーゼ染色
・ブロッブ間領域細胞
高空間周波数選択性
・方向選択性細胞
縞刺激を縞と直角に動かすと周期的応答(単純型細胞)
視覚刺激なくても発達(Rochefort,Konnerth,2011)
・両眼性細胞
・左眼優位型細胞
・右眼優位型細胞
(Hubel&Wiesel,1962)
○単純型細胞(simple cell)楕円形受容野
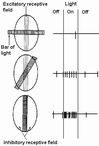
特定の位置の特定の方位の線分やエッジの検出
生成機構をLGNdの神経との同時記録で推測(Roid&Alonso,1995,nature)
1) サンドイッチON中心型
2) サンドイッチOFF中心型
3) サンドイッチずれてる型
4) ON左OFF右型
・両眼から入力を受ける細胞がある
○複雑型細胞(complex cell)矩形受容野

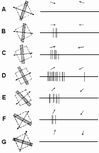
位置にあまり関わらず特定方位のエッジを検出
方向選択性(受容野の長軸と直角に片方の方向)
○超複雑型細胞(hypercomplex)矩形受容野

終端抑制型細胞(end-inhibition cell)
特定の長さの線分や端点検出
・両眼視差に応答する神経細胞
・面に対する応答(全細胞の15%くらい)
明るさの対比効果が見られた(Kinoshita&Komatsu,2001)
・type1:周辺の影響なし
・type2:周辺によって抑制される
・type3:周辺によって促進される
周辺の影響の潜時が長い(Rossi,Paradiso,1996)
数Hzのちらつきで周辺の誘導効果が消失した
horizontalな神経伝達は時間がかかるようだ
・方向選択性細胞
(Anderson,Ferster,2000; Monier,Fregnac,2003)
【特徴】
◆網膜部位再現(topographic mapping, retinotopic mapping)
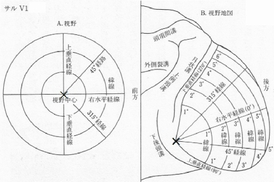
・外側膝状体背側核同様
・烏距溝(calcarine fissure)の上側(18野)が網膜の上側(視野下部)
・下側(17野)が下側(視野上部)
・網膜偏心度が小さいほど視覚分解能が高い
・地図の決定には水平経線・垂直経線の刺激を用いる(Hadjikhani,1998)
(この刺激への応答でV1d,V2d,V3d,V3A,V4dの境界線決定)
◆後細胞への出力
・1細胞が異なる時空間情報を異なる領野へ送る
(mice: Glickfeld,Reid,2013)
◆臨界期
・生後25〜27日齢に4日間眼帯で対側V1の細胞数の偏りがなくなる
GABA合成酵素欠損マウスではこの効果消失(Hensch et al.,1998)
・NMDA受容体の長いgating時間が数十日で短くなる
(rat 4層: Carmignoto&Vicini,1992)
◆統合
・異なる方位選択性入力が樹状突起上に散在
(Jia,Konnerth,2009)
・類似の方位選択性の細胞同士は結合率高い(Lee,Reid,2016)
◆視覚入力でシナプス入力が非同期に(Tan,Priebe,2014)
2014/05/13 masashi tanaka
